Wir alle lernen. Jeden Tag und überall: In Bibliotheken, Hörsälen, Laboren, im Bett oder fünf Minuten vor der nächsten Klausur: Aber wie funktioniert das eigentlich?
Lernprozesse finden im Gehirn statt. Dieses besteht aus Nerven, die über Synapsen miteinander in Verbindung stehen. Um etwas über die Informationsaufnahme und Speicherung zu erfahren, müssen wir uns also zuerst mit der Funktionsweise und Kommunikation von Nerven auseinandersetzen.
Eine Nervenzelle kann man sich vorstellen wie eine Disco. Begrenzt wird sie von einer Plasmamembran, die aus einer Schicht von ungeladenen Fettmolekülen besteht und verschiedene Funktionen ausübt. Die Membran hält die Zelle zusammen und sorgt dafür, dass wichtige Moleküle und Teilchen nicht unkontrolliert in die Zelle gelangen oder sie verlassen können. Sie ist selektiv permeabel, das heißt sie besitzt Kanäle, die wie Türsteher dafür sorgen, dass ausschließlich in die Zelle gelangt, was auch hinein soll – also nur bestimmte Teilchen unter bestimmten Bedingungen.
Zwischen dem Innenraum der Zelle und ihrer Umgebung herrschen große Unterschiede in der Konzentration einzelner Ionen (elektrisch geladene Teilchen). Im Ruhezustand gibt es in der Zelle (intrazellulär) eine große Zahl von einfach positiv geladenen Kaliumionen (K+) und negativ geladenen organischen Teilchen, wohingegen außerhalb der Zelle (extrazellulär) eine Anreicherung von einfach positiv geladenen Natriumionen (Na+) und einfach negativ geladenen Chloridionen (Cl–) sowie zweifach positiv geladenen Calciumionen (Ca2+) herrscht. Die Plasmamembran wirkt hierbei als Barriere, ihre selektive Durchlässigkeit verhindert, dass es auf Grund der Brownschen Molekularbewegung zu einem Gleichgewicht (Konzentrationsausgleich) der Teilchen intra- und extrazellulär kommt.
Brownsche Molekularbewegung
Durch die sogenannte „Wärmebewegung“ stoßen Wassermoleküle ständig ungeordnet in elastischen Stößen gegeneinander.
Diffusion entsteht dadurch, dass Teilchen an Orten, in denen sie in hoher Konzentration vorliegen, häufiger aneinanderstoßen, als in Bereichen, in denen sie in niedriger Konzentration vorliegen und sich dadurch gleichmäßig in einem Raum verteilen. Die Höhe des Unterschieds der Konzentration in zwei Regionen (Konzentrationsgradient) ist dabei die treibende Kraft.
Da Wärme ein Maß für die mittlere kinetische Energie (Bewegungsenergie) von Teilchen ist, gibt es in flüssigem Wasser mäßige Molekularbewegung, wohingegen Eis kalt und fest ist (kaum Molekularbewegung) und heißes Wasser verdampft (viel Molekularbewegung). Folglich laufen auch Diffusionsprozesse unter warme Bedingungen schneller ab.
Zwischen Intra- und extrazellulärem Raum gibt es also zwei Gefälle:
- Den Konzentrationsunterschied von Natrium-, Kalium-, Chlorid- und Calciumionen
- Den Ladungsunterschied, denn extrazellulär gibt es mehr positive Ladungen als intrazellulär.
Bedingt durch die Ladungstrennung und die unterschiedlichen Ionenkonzentrationen entsteht das Ruhemembranpotential von etwa -70mV (milliVolt), die Potentialdifferenz der Ladungen intra- und extrazellulär.
Nerven leiten Reize über kurze elektrische Signale weiter, die sogenannten Aktionspotentiale, bei denen das Membranpotential durch den Einstrom von Natriumionen (Na+) kurz positiv wird (Depolarisation). Die Natriumionen strömen in die Zelle, weil sie durch das Konzentrationsgefälle sowie die intrazelluläre negative Ladung angezogen werden. Das Aktionspotential wird über die Membran durch unterschiedliche ionenspezifische Kanäle entlang des Nervs und über Synapsen von einem zum anderen Nerv transportiert.
Eine Synapse besteht aus zwei Teilen: Erstens dem Endknöpfchen (Ende des Nervs, welcher das Signal weiterleitet), zweitens der postsynaptischen Membran (Stück der Membran des Nervs, der das Signal empfängt), die direkt unter dem Endknöpfchen liegt. Zwischen Endknöpfchen und postsynaptischer Membran befindet sich der synaptische Spalt, eine Lücke von ungefähr 20nm Breite, die von Transmittern überwunden werden muss. Transmitter sind Botenstoffe, die ein Signal von einem Nerv auf einen anderen übertragen. Im menschlichen Gehirn handelt es sich dabei meist um Glutamat, eine Aminosäure. Die Transmitterausschüttung durch das Endknöpfchen wird von einem Aktionspotential ausgelöst, welches dort ankommt und über die Synapse übertragen werden soll. Auf der anderen Seite, an der postsynaptischen Membran, bindet der Transmitter an Ionenkanäle, die sich daraufhin öffnen und im zweiten Nerv ein Aktionspotential auslösen. An einem Nerv befinden sich, zur Vernetzung und Verrechnung, häufig mehrere Synapsen.
Nochmal bildlich: Tagsüber im Ruhezustand ist die Stimmung im Club ziemlich negativ (-70mV), keiner tanzt und missmutige Mitarbeiter (intrazellulär hohe Konzentration von K+ und organischen Anionen) räumen auf. Gegen Abend beginnen sich die Feierwütigen (extrazellulär hohe Konzentration von Cl–, Na+ und Ca2+) vor der Tür zur sammeln, aber die Türsteher (Ionenkanäle) lassen nur die „coolen Leute“ (Na+) mit der richtigen Einstellung herein, damit die Stimmung positiv (Depolarisation) wird und die Party steigen kann (Aktionspotential). Zwischen den Clubs (zwei Nerven) bewegen sich die Gäste mit Hilfe von Bussen (Transmittern). Das funktioniert nur an bestimmten Haltestellen (Synapse = Endknöpfchen + postsynaptische Membran). Manche Discos sind besser angebunden als andere und besitzen mehrere Haltestellen (mehrere Synapsen an einer Nervenzelle).
Und was hat das jetzt mit Lernprozessen zu tun?
Die Nervenzellen des Gehirns sind veränderlich, können sich neu untereinander verknüpfen und sich, ähnlich wie Muskeln, an Belastungen anpassen. Man spricht dabei von „Plastizität“. Unter „Lernen“ versteht man dabei, dass Nervenzellen bestimmte Reize verstärkt oder in eine bestimmte Richtung weiterleiten. Ein anerkanntes Modell zur Erklärung des Lernprozesses ist die Langzeitpotenzierung, dabei handelt es sich um eine langfristig verstärkte Weiterleitung bestimmter Reize durch eine Nervenzelle, bedingt durch einen außergewöhnlich großen, oft wiederholten Reiz oder durch zwei Reize gleichzeitig. Die Langzeitpotenzierung beruht auf zwei Typen von Ionenkanälen:
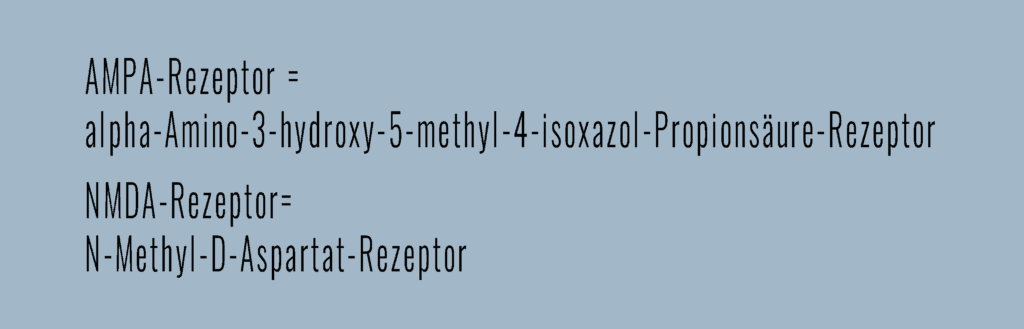
- AMPA-Rezeptor: Der AMPA-Rezeptor befindet sich in der postsynaptischen Membran und hat eine Bindungsstelle für den Transmitter Glutamat. Wird Glutamat vom Endknöpfchen als Zeichen für ein eingelaufenes Aktionspotential ausgeschüttet, so bindet es an den AMPA-Rezeptor, welcher daraufhin seine Struktur ändert und für Natrium durchlässig wird. Positive Natriumionen strömen aus dem Extrazellulärraum in den Nerv ein, das Innere der Nervenzelle wird kurzfristig positiv geladen, ein Aktionspotential ausgelöst und weitergeleitet.
- NMDA-Rezeptor: Das Grundprinzip des NMDA-Rezeptors ist dem des AMPA-Rezeptors sehr ähnlich. Auch er besitzt eine Bindungsstelle für Glutamat und ist permeabel für Natrium. Im Normalfall ist seine Pore, die Durchtrittsstelle für die Ionen, jedoch durch ein zweifach positives Magnesiumion (Mg2+) verschlossen. Das heißt, auch wenn Glutamat an den NMDA-Rezeptor bindet, können keine Ionen den Kanal passieren. Damit die Pore durchlässig wird, muss ein Aktionspotential vorangegangen und die Nervenzelle schon leicht positiv geladen (vordepolarisiert) sein. Dann verlässt das Mg2+ die Pore, abgestoßen durch die positive Ladung der Zelle. Folgt nun ein zweites Aktionspotential, schüttet das Endknöpfchen erneut Glutamat aus. Das bindet an den NMDA-Rezeptor, dessen nun nicht mehr verstopfte Pore öffnet sich und lässt Natrium- und Calcium-Ionen, die dem Konzentrationsgradienten nach intrazellulär folgen, passieren. Die Vordepolarisation kann entweder durch eine Synapse, die kurz hintereinander mehrere Signale weiterleitet oder durch mehrere Synapsen an einem Nerv, die Aktionspotentiale gleichzeitig weitergeben, geschehen.
Neben Natriumionen können auch Calciumionen den NMDA-Rezeptor passieren. Calciumionen spielen eine sehr wichtige Rolle, denn sie bewirken Umbauarbeiten in der Zelle, und sorgen so unter anderem dafür, dass Signale verstärkt weitergeleitet werden.
Informationsaufnahme an Rezeptoren: Wie gelangen die Informationen aus der Umwelt denn eigentlich ins Gehirn?
Die Reizaufnahme, auch Rezeption genannt, beginnt immer mit einem Rezeptor. Es gibt zahlreiche Rezeptoren, von den Photorezeptoren des Auges, über Nozizeptoren (Schmerzrezeptoren) auf der Haut, bis hin zu den Ruffinikörperchen der Gelenke, die uns ständig mitteilen, in welche Richtung unsere Gelenke gerade gebogen sind.
Das Funktionsprinzip der Rezeptoren ist stets gleich: Ein Reiz führt zur Öffnung von Ionenkanälen, sodass ein Aktionspotential entsteht, welches aus der Peripherie zum Gehirn weitergeleitet wird (Transmission). Wie der Reiz die Ionenkanäle öffnet, ist jedoch verschieden und immer abhängig von der Art des Reizes. So öffnen sich zum Beispiel in Mechanorezeptoren (Rezeptoren, die Berührungen wahrnehmen) die Ionenkanäle durch Druck, in olfaktorischen Rezeptoren (Geruchsrezeptoren) durch die Bindung bestimmter Moleküle und im Innenohr versetzen Schwallwellen kleine Härchen in Schwingungen. Die „Übersetzung“ eines Reizes in die Änderung eines Membranpotentials, sodass ein Aktionspotential entsteht, wird als Transduktion bezeichnet. Erreicht das Aktionspotential nach erfolgreicher Transmission das zentrale Nervensystem und wird dort zu einem Eindruck (süß, sauer, heiß, kalt etc.) verarbeitet, so spricht man von Perzeption, der Wahrnehmung.
Zum Ablauf der Langzeitpotenzierung: Wird eine Synapse besonders hochfrequent genutzt, also Aktionspotentiale kurz hintereinander weitergeleitet oder leiten mehrere Synapsen gleichzeitig Aktionspotentiale an einen Nerv weiter, kommt es häufiger zur Aktivität der NMDA-Rezeptoren. In der Folge erhöht sich die Calcium-Konzentration in der Zelle und setzt Umbaumechanismen in Gang, die, etwa durch den Einbau von weiteren Ionenkanälen oder Bildung neuer Synapsen, dafür sorgen, dass künftig eintreffende Signale vor ihrer Weiterleitung verstärkt werden. Die Sensibilisierung der Nervenzelle kann Tage bis Wochen anhalten und ist der Grund dafür, warum wir Dinge miteinander assoziieren können.
Bildlich gesprochen: Der AMPA-Rezeptor ist ein ziemlich durchschnittlicher Türsteher, einfach gestrickt und sehr zuverlässig wartet er auf den Bus (Ausschüttung des Transmitters Glutamat bei Aktionspotential) und lässt die Gäste (Na+) hinein. Der NMDA-Rezeptor ist von der fauleren Sorte, er stellt einen Stuhl unter die Türklinke (Mg2+) und geht schlafen. Im Normalfall passiert dann gar nichts, die Party läuft und der Türsteher bekommt seinen Schlaf. Manchmal jedoch strömen so viele Gäste in den Club (vorangegangene Aktionspotentiale > Vordepolarisation), dass die Hitze darin nicht auszuhalten ist und jemand auf der Suche nach frischer Luft den Stuhl wegschiebt, sodass die Tür aufgeht. Jetzt gibt es kein Halten mehr, mit normalen Gästen (Na+) strömen auch Krawallmacher (Ca2+) in den Club und beginnen ihn auseinanderzunehmen. An diese Randale erinnert man sich noch lange und als Reaktion darauf werden bei der nächsten Party mehr Türsteher (Ionenkanäle) eingesetzt und bei fortdauerndem Besucherandrang neu Bushaltestellen (Synapsen) gebaut. Aus einer starken Erfahrung in Folge eines großen Besucherandrangs (mehrere Aktionspotentiale), wurde gelernt.
Weiterführende Literatur
Pape, H-C. et al. (1979): Physiologie. Stuttgart: Thieme. 7. Auflage 2014.
Reece, J. et al. (2016): Campell Biologie. Hallbergmoos: Pearson. 10.Auflage 2016.
Schmidt, R. et al. (1936): Physiologie des Menschen mit Pathophysiologie. Heidelberg: Springer. 31. Auflage 2010.
Tipler, P. et al. (1994): Physics for Scientists and Engineers. Heidelberg: Springer. 7. Auflage 2015.